(O texto a seguir é a tradução a partir do inglês deste PDF)
Os animais se curam, mas as árvores se compartimentam. Elas suportam uma vida inteira de ferimentos e infecções, estabelecendo barreiras internas que resistem à propagação dos microrganismos invasores.
por Alex L. Shigo (1985)
As árvores têm um recorde espetacular de sobrevivência. Durante um período de mais de 400 milhões de anos, elas evoluíram como os organismos mais altos, mais maciços e mais longevos que já habitaram a terra. No entanto, às árvores falta um meio de defesa que quase todos os animais possuem: elas não podem se afastar das forças destrutivas. Por não poderem se mover, todos os tipos de inimigos vivos e não vivos – fogo, tempestades, microrganismos, insetos, outros animais e mais tarde o ser humano – causaram-lhes ferimentos ao longo de sua história. As árvores sobreviveram porque sua evolução as transformou em organismos altamente compartimentados; isto é, elas separam com barreiras a madeira ferida e infectada.
Nesse aspecto, as árvores são radicalmente diferentes dos animais. Fundamentalmente, os animais se curam: eles preservam sua vida fazendo bilhões de reparos, colocando novas células ou células rejuvenescidas nas posições das antigas. As árvores não podem curar; elas não fazem reparos. Em vez disso, elas se defendem das consequências de ferimentos e infecções por meio de barreiras de proteção contra os danos. Em resumo, elas se compartimentam. Ao mesmo tempo, elas colocam novas células em novas posições; com efeito, elas crescem uma nova árvore sobre a antiga a cada ano. Os resultados mais óbvios do processo são os anéis de crescimento, que são visíveis na seção transversal de um tronco, uma raiz ou um ramo.
As defesas que as árvores usam sugerem uma nova visão da sua biologia, na qual o papel da patologia das árvores ganha destaque. Elas têm sido guiadas através da evolução por sua necessidade de se defenderem contra ataques enquanto se mantêm plantadas no chão.
Para compreender as doenças e a deterioração das árvores é essencial compreender o funcionamento e o crescimento normal delas. As árvores, juntamente com as gramíneas e algas, são os maiores capturadores de energia da Terra. Em particular, as florestas, que cobrem cerca de um décimo da superfície do planeta, capturam cerca da metade de toda a energia que entra na biosfera. A energia, que entra como radiação solar, serve para transformar dióxido de carbono e água em carboidratos, a forma química na qual a energia é armazenada. Por sua vez, os carboidratos impulsionam o crescimento, a manutenção, a reprodução e a defesa delas.
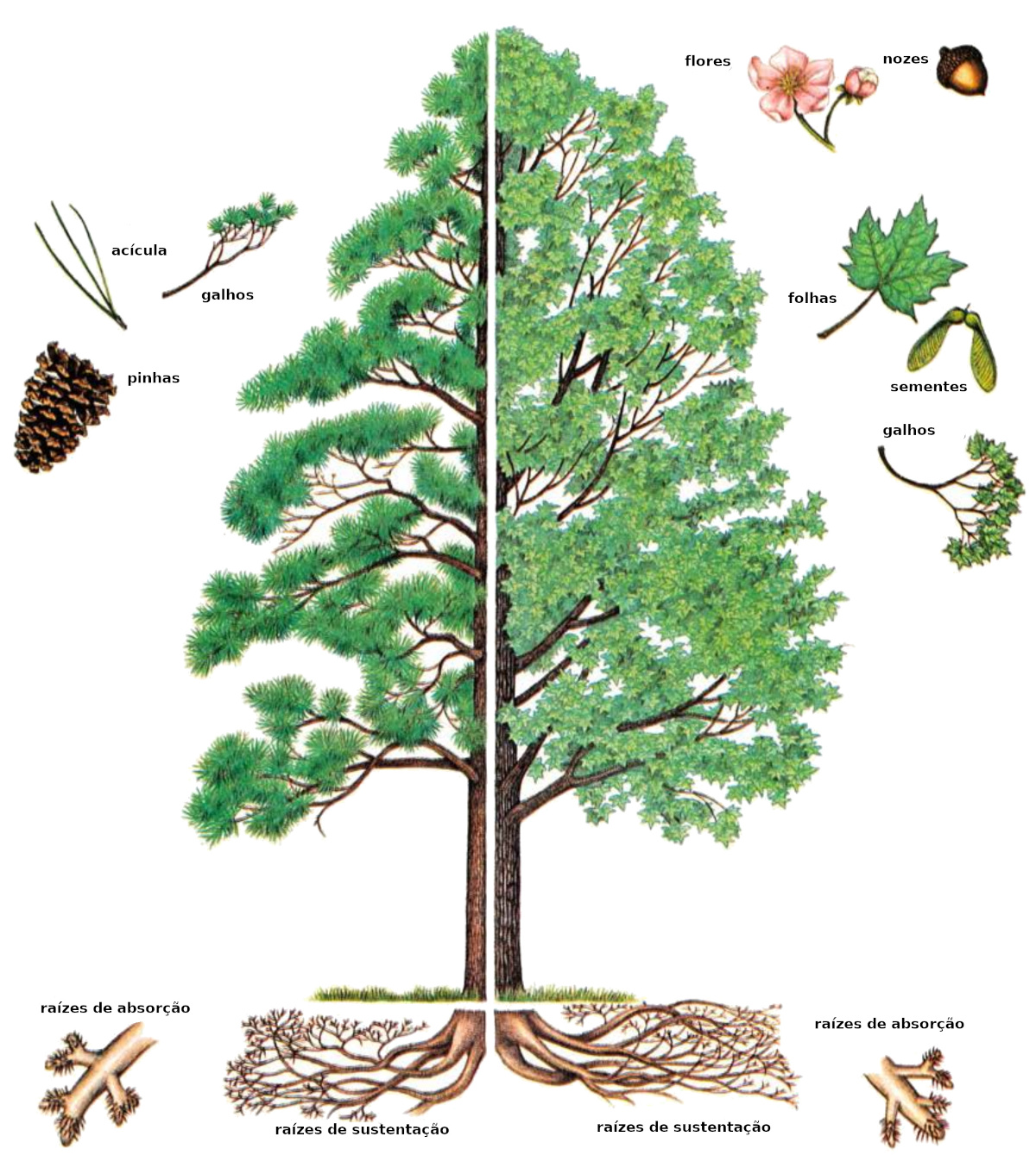
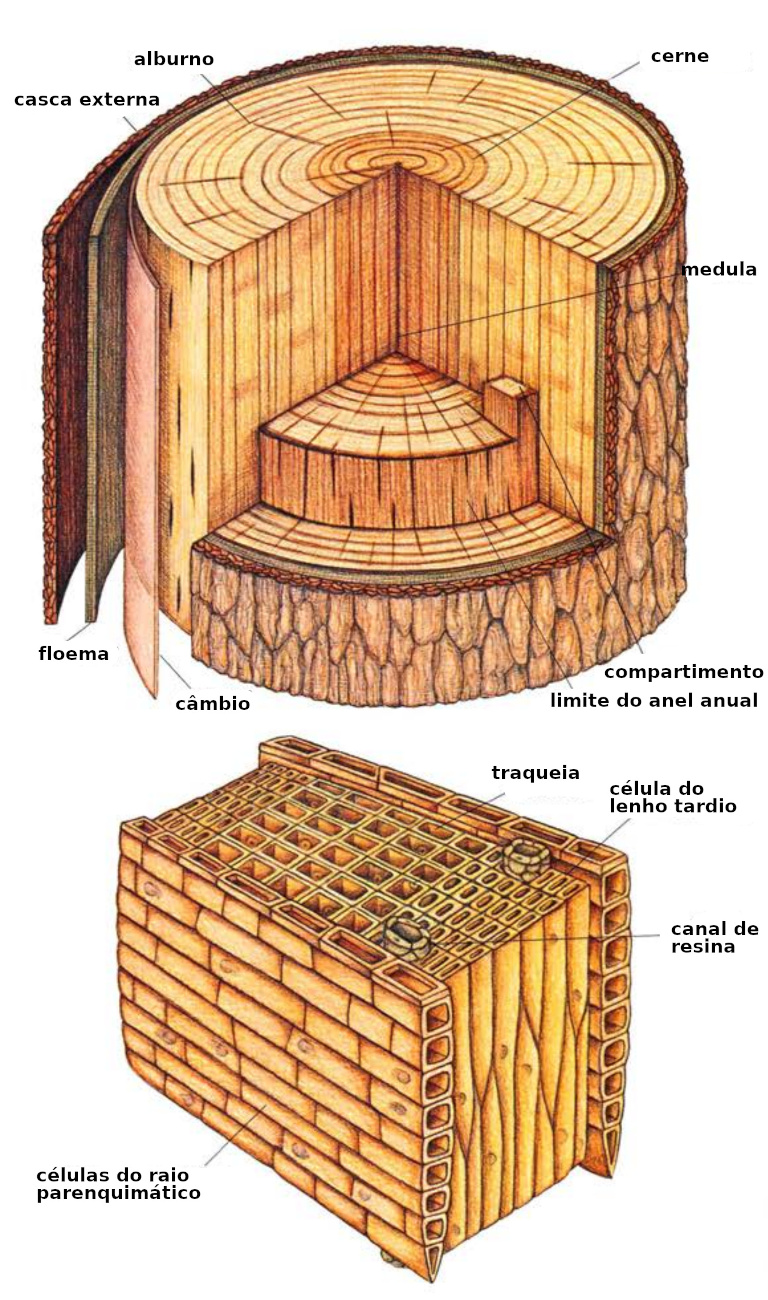
As próprias árvores – isto é, as coníferas e as madeiras duras (as angiospermas e gimnospermas) – são plantas perenes, lenhosas, compartimentadas, que separam pedaços internos de si. Além disso, a maioria das árvores são longevas, maciças e altas. Sua estrutura interna segue um padrão característico. A geração de novas células na árvore é a função do câmbio vascular, uma fina camada cilíndrica encontrada no tronco, nas raízes e nos galhos. Na direção externa, o câmbio estabelece o floema, ou casca interna, a camada que transporta líquidos para baixo – especificamente, as substâncias produzidas pela fotossíntese nas folhas. Na direção interna, o câmbio se assenta sobre camadas concêntricas do xilema, ou simplesmente o lenho, que transporta água e substâncias solúveis em água para cima.
A produção de células internas merece um exame minucioso. Em uma escala ampla, o câmbio estabelece uma camada interna a cada ano; estes são os anéis de crescimento anual, destacados em uma seção transversal da árvore. (Em árvores tropicais, entretanto, os anéis de crescimento são indistintos.) Vista em uma escala mais detalhada, o câmbio forma dois tipos básicos de células: aquelas com seu longo eixo perpendicular ao eixo do tronco, de uma raiz ou de um ramo, e aquelas com seu longo eixo paralelo ao eixo do tronco, raiz ou ramo. As células perpendiculares tornam-se parênquima radial, que formam divisórias radiais na madeira. Por outro lado, as células longitudinais preenchem os compartimentos entre os raios.
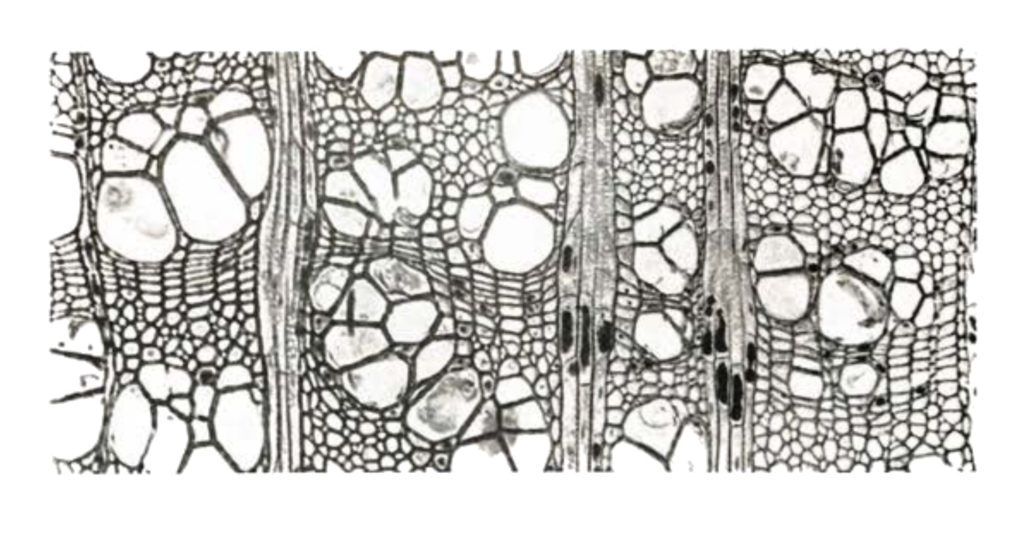
As células longitudinais são de três variedades. Em algumas, o conteúdo vivo morre em poucos dias ou semanas, deixando apenas uma parede celular tubular. Em madeiras duras, tais células são chamadas de vasos; em coníferas são chamadas de traqueias. Elas servem para o transporte de líquidos. Em outras, a parede celular é espessa. Tais células, chamadas células de fibra ou traqueias de fibra, fornecem suporte mecânico para a madeira. Finalmente, as células chamadas parênquima retêm seu conteúdo vivo atrás de uma parede celular fina. (Em contraste, vasos e traqueias têm um interior oco sob uma parede celular espessa). Células parenquimatosas armazenam nutrientes e outros materiais. Nelas, o citoplasma pode permanecer vivo por anos. Às vezes, por mais de um século. O complexo intertravamento das várias células numa madeira combina com as próprias paredes celulares resistentes para dar força ao seu tecido. O desenho de intertravamento continua na construção molecular das paredes celulares, e até mesmo nas moléculas individuais da parede celular: celulose e lignina.

Em qualquer momento da vida da árvore, as camadas mais recentes do lenho, aquelas em que o parênquima ainda retém seu conteúdo vivo, formam o alburno. Em muitas delas, as camadas mais antigas, mais próximas do eixo da árvore, formam uma região chamada de cerne, que muitas vezes é de cor mais escura. Parte desse teor escuro se deve ao depósito de substâncias conhecidas comumente como extrativas. O cerne tem um alto grau de autossustentação mecânica; assim, ele continua servindo à árvore. Por outro lado, o cerne não tem capacidade de armazenar nutrientes e transportar substâncias.
O estudo da anatomia das árvores sugere uma série de maneiras pelas quais elas podem ser vistas conceitualmente. Em primeiro lugar, as árvores são geradoras de tecidos. Em essência, a germinação de uma semente é a ativação de um gerador de células. Ela tem a capacidade de proliferar células, mas sempre em novos locais; a árvore não tem capacidade de re-armazenar ou regenerar tecidos que já estão alocados. O gerador é o câmbio. Em segundo lugar, a madeira é um arranjo altamente ordenado de diferentes tipos de células em diferentes estágios de envelhecimento. Em terceiro lugar, uma árvore é uma hierarquia de compartimentos. Dentro do tronco, das raízes e dos galhos, os maiores compartimentos são os anéis anuais. Depois, vêm os grupos de células em cada anel, compartimentados por raios, e em seguida as células individuais.
O estudo de como as árvores respondem a ferimentos ou infecções sugere um conceito adicional. As árvores respondem compartimentando-se: elas tentam separar a região ferida ou infectada. Elas não matam nem impedem a atividade dos microrganismos nos compartimentos que são murados. Nem respondem de maneiras específicas a microrganismos específicos; a compartimentação vem em resposta ao fato do ferimento.
Em linhas gerais, a árvore responde de três formas a ferimentos e infecções. Na primeira delas, os limites dos compartimentos já existentes são reforçados para resistir à propagação da deterioração. Na maior parte dos casos, o endurecimento é conseguido por meios químicos. No alburno, o metabolismo das células parenquimatosas vivas muda, de forma a alterar o conteúdo das células. No alburno, as reações enzimáticas ocorrem no tecido, que já não está mais vivo.
Os detalhes ainda são mal compreendidos. Mesmo assim, em circunstâncias normais, as células do lenho dedicam seus caminhos bioquímicos ao armazenamento de energia química sob a forma de carboidratos. Normalmente, as moléculas estão em forma reduzida: seu conteúdo de elétrons é relativamente grande. Após a lesão da madeira, a atividade bioquímica nas células ao redor da lesão é desviada para novos caminhos metabólicos, de modo que as moléculas tendem a ser oxidadas; ou seja, prótons, ou íons de hidrogênio, são fixados. Numa escala mais ampla, o conteúdo celular passa por um processo químico muito semelhante ao curtimento do couro. Assim, surgem moléculas como o ácido gálico e o ácido tânico. Elas compartilham a propriedade de serem ricas em fenol (seis anéis de carbono contendo grupos de hidroxilas, ou OH). Os fenóis ocupam o interior das células; eles também impregnam as paredes celulares. Os fenóis descolorem a madeira em tons de vermelho, verde ou azul, dependendo dos detalhes das vias químicas, que são determinadas pela genética de uma determinada espécie de árvore. Mais importante para as defesas da árvore, os compostos de fenol tendem a ser antimicrobianos.
Na segunda resposta que dá à lesão e/ou infecção, a árvore cria uma nova parede por meios anatômicos e químicos. Primeiro, o câmbio muda o padrão pelo qual gera novas células. Após a lesão, poucas células de canais são produzidas. As células de fibra também são produzidas em menor quantidade. Por outro lado, o parênquima, ou células que retêm seu conteúdo vivo, são produzidas em maior quantidade. Agora, porém, elas são menores, e sua atividade metabólica é alterada para que seu conteúdo químico resista aos microrganismos. A nova parede, ou zona de barreira, é a causa de muitos dos defeitos encontrados nos produtos madeireiros. Por exemplo, ela pode fazer com que o lenho na árvore viva se separe ao longo de um círculo. O defeito é conhecido como greta.

A terceira resposta que a árvore dá é continuar crescendo. As árvores sobrevivem após ferimentos e infecções se tiverem tempo, energia e capacidade genética suficientes para reconhecer e compartimentar o tecido ferido e infectado enquanto geram o novo tecido que irá manter sua vida. Em certa medida, portanto, uma árvore ferida ou infectada se assemelha a um navio fortemente compartimentado ou a um velho submarino sob ataque. Quando o navio é atingido por um torpedo, a tripulação se apressa para selar a área danificada. Quanto mais rapidamente for a ação da tripulação e quanto mais fortes forem as paredes circunscrevendo o compartimento danificado, menor será a extensão do dano. Mas após o dano ser contido, o compartimento ou compartimentos danificados não são mais acessíveis. Neste ponto, a analogia termina. A árvore sobrevive crescendo o que equivale a uma nova árvore (com um novo conjunto de compartimentos) sobre si mesma durante a próxima estação de crescimento.
É preciso dizer que as zonas de reação da árvore (seus limites quimicamente reforçados) não são absolutas: pode ser que elas recuem, rápida ou lentamente, da infecção, pois certos microrganismos superam a defesa química. Deve-se dizer também que os fenóis são venenosos não só para os microrganismos, mas também para a árvore. Com efeito, ela envenena parte de si mesma na tentativa de estancar uma invasão. Ela sobrevive porque ao mesmo tempo em que fortalece as barreiras, também está criando uma nova árvore.
Notavelmente, a capacidade de uma árvore de separar partes de si mesma é muito parecida com a resposta que dá a ferimentos e infecções. Ou seja, a separação é um aspecto da compartimentação. Em particular, acículas, folhas, estruturas reprodutivas e raízes absorventes que cumpriram seu programa genético são separadas da árvore por paredes (raízes absorventes são as raízes finas, não lenhosas, que absorvem substâncias do solo). Ramos, galhos e raízes grandes também podem ser murados, digamos na sequência de uma lesão ou infecção, ou após o ramo, galho ou raiz ter atingido um determinado estágio de senescência. (É difícil saber exatamente em que estágio; a programação genética de envelhecimento nas árvores é mal compreendida). O que acaba de ser dito exige uma qualificação: as árvores não removem ativamente partes de si mesmas. Vento, neve, gelo, animais e outros agentes fazem com que as partes muradas caiam da árvore. Enquanto isso, as raízes absorventes que foram muradas são digeridas pelos microrganismos do solo.
Dirijo-me agora para o outro lado da briga entre a árvore e seus inimigos. Começarei com um ferimento novo realizado, digamos, por um animal. A ferida fornece novo espaço e nutrientes para uma série de organismos invasores, incluindo insetos, nematóides, bactérias e fungos. Enquanto competem entre si pelo novo espaço e nutrientes, as células vivas no alburno subjacente à ferida estão reagindo à invasão, passando por uma variedade de mudanças bioquímicas que levam à produção de defesas químicas à base de fenol.
Os microrganismos atacam as feridas das árvores de diversas maneiras. Certas bactérias e fungos infectam a casca interna e ficam lá, criando as doenças conhecidas como cancros anuais. Outros microrganismos invadem as feridas e permanecem no tecido do alburno ferido, criando as chamadas feridas de podridão. Ainda outros microrganismos infectam a casca interna, se estabelecem e depois infectam o lenho. Através da repetição anual do processo, eles criam cancros perenes. Finalmente, alguns microrganismos atacam uma ferida, primeiro infectando o alburno e depois infectando a casca interna. Novamente, o processo se repete a cada estação de crescimento. Estes são os chamados fungos da podridão seca. Eles são notavelmente insidiosos. Quando a podridão do cancro evoluiu do lenho para a casca, ela forma um tecido duro, semelhante a uma cunha, que mata o câmbio subjacente. A árvore reage reativando suas defesas compartimentadoras. O fungo, por sua vez, invade a nova ferida, a partir da qual cresce outra cunha. O movimento de vai e vem pode continuar até que a árvore seja cingida.
Os primeiros microrganismos a terem sucesso na invasão da árvore são chamados de pioneiros. Eles podem simplesmente ser capazes de tolerar as alterações químicas provocadas na madeira pelo ferimento da árvore. Em alguns casos, porém, sua constituição genética os torna capazes de digerir as defesas químicas. Tipicamente, mas nem sempre, os pioneiros são bactérias, juntamente com certas espécies de fungos. Entre estas últimas se destacam os Himenomicetos, que causam deterioração nas árvores, e os Deuteromicetos e Ascomicetos, que em sua maioria não causam a deterioração em si.
Um ponto crucial sobre a infecção é que os microrganismos se estabelecem em uma sequência particular. Quando os microrganismos pioneiros superam os químicos inibidores, eles abrem o caminho para outros invasores, aqueles que teriam sucumbido às defesas da árvore. Assim, a invasão toma a forma de uma sucessão de organismos, um padrão essencial para a sobrevivência dos invasores. Evidentemente, nenhum microrganismo “come veneno” para ajudar seus sucessores. Cada organismo age de forma a aumentar sua conquista de espaço e energia. Portanto, alguns dos pioneiros podem realmente criar ou preservar condições que inibem a infecção por fungos agressivos que degradam a madeira. Tais pioneiros podem, em última instância, ser a base para o controle biológico do apodrecimento das árvores. Estudos de certos fungos, tais como as espécies de Trichoderma, já mostram que os fungos prosperam no alburno alterado por feridas, mas não desintoxicam os produtos químicos que mantêm fora os fungos causadores do apodrecimento.
Suponha que uma sucessão de microrganismos invasores seja bem sucedida na digestão do alburno alterado pela ferida. O sucesso deles não necessariamente condena a árvore. Por um lado, ela está gerando novas células ao redor das antigas. Se a árvore puder gerar novas células mais rapidamente do que as antigas estão sendo digeridas, ela tem uma boa chance de sobrevivência. A morte de uma árvore, se ela acontecer, pode ser mecânica ou biológica. Ela morrerá se o tronco quebrar, por exemplo. Alternativamente, ela morrerá se o câmbio – o gerador de células – for morto. Além disso, morrerá se grande parte do seu tecido tiver sido murada durante uma vida inteira de defesas contra ferimentos e infecções de forma que os compartimentos restantes são insuficientes para armazenar a energia da árvore.
O novo entendimento sobre as árvores como organismos compartimentadores não surgiu de uma só vez. Na verdade, ele veio como uma contradição de noções anteriores, algumas das quais foram desenvolvidas logo após os fundamentos da biologia moderna terem sido estabelecidos há um século. Parece uma coisa banal de se dizer, mas as árvores são fundamentalmente diferentes dos animais, e muito do fracasso em compreendê-las deriva de confundir inconscientemente os dois.
Antes de 1845, a explicação mais aceita sobre como a vida se dá era que ela se origina espontaneamente (ou seja, por geração espontânea) a partir de material inorgânico. Sabia-se alguma coisa sobre microrganismos; em particular, já haviam se dado conta da associação entre madeira apodrecida e fungos. A ideia, entretanto, era que a decomposição dá origem aos fungos. Na esteira de Heinrich Anton De Bary, Louis Pasteur e do desenvolvimento da teoria dos germes, que atribui doenças a organismos minúsculos e prejudiciais, o patologista alemão Robert Hartig reverteu a ideia ao propor que os fungos dão origem à decomposição. Hartig mostrou que os esporóforos, ou corpos frutíferos, encontrados nas feridas das árvores, e os micélios, ou organismos em forma de filamentos, encontrados na madeira apodrecida, representam diferentes estágios de vida de um mesmo organismo fúngico. A observação, e a inversão da hipótese, preparou o terreno para a ciência da patologia das árvores.
Estudos subsequentes sobre deterioração foram feitos por investigadores interessados principalmente no estrago de madeiras serradas. Assim, os estudos se basearam na remoção do lenho das árvores, seguida pela investigação laboratorial das mudanças no tecido. Os estudos se centraram no cerne, ou em qualquer caso, na madeira mais escura que o alburno. A estratégia por trás dos estudos tinha como justificativa o fato de que o cerne de uma árvore está morto, ou seja, ele é um tecido não responsivo. Até certo ponto, os estudos foram bem sucedidos: químicos e patologistas passaram a entender como os fungos (em particular, suas enzimas) digerem a madeira. Ainda assim, os processos na árvore viva não foram considerados. Embora o cerne esteja morto pelos padrões animais, ele é reativo a ferimentos e infecções. Alguns textos ainda afirmam que a deterioração nas árvores não pode ser considerada uma doença porque apenas o cerne morto é infectado. A deterioração é, na verdade, a principal doença de todas as árvores no mundo.
Os primeiros pesquisadores não foram capazes, portanto, de reconhecer as defesas ativadas por ferimentos e infecções, que alteram a madeira quando a árvore é ferida. Em uma árvore viva, a maioria das feridas não produzem deterioração porque os microrganismos que infectam a ferida se deparam com a madeira que mudou. Em contraste, os fungos que produzem o apodrecimento que ataca a madeira serrada não enfrentam a força contrária dela. Os microrganismos simplesmente competem entre si.
A tecnologia crucial que permitiu aos pesquisadores desenvolver uma nova compreensão da deterioração nas árvores foi a motosserra. Nos anos 1940, o equipamento era suficientemente potente para que a pessoa que o operasse na floresta pudesse facilmente cortar seções longitudinais através das árvores, expondo colunas de madeira descolorida e em decomposição sob os anéis de crescimento que a árvore tinha desenvolvido após a lesão. Antes de 1940, as árvores já haviam sido dissecadas, mas a maioria dos cortes eram transversais. Apenas alguns poucos investigadores, como o patologista George H. Hepting, que trabalhava no delta do Mississípi nos anos 1930, procederam de outra forma. Hepting, usando serras e eixos transversais, pôde ver o que outros viram mais tarde: que em longos trechos de árvores, a madeira gerada após uma ferida não foi invadida pelos fungos que infectaram a ferida em si.
Nos anos 1960 e 1970, eu mesmo tive a oportunidade de dissecar milhares de árvores, primeiro no nordeste dos EUA e depois na Europa, Índia, Porto Rico e Austrália. Fiquei impressionado, por um lado, com os padrões ordenados de descoloração e, por outro, com a sucessão ordenada dos microrganismos invasores. Meus colegas e eu fizemos então experiências com árvores vivas. Ainda mais tarde, estudos bioquímicos foram realizados por Walter C. Shortle, meu colega do Serviço Florestal do Departamento de Agricultura dos Estados Unidos em Durham , N.H.
Em um esforço para definir um ponto em comum entre as respostas que uma árvore dá a ferimentos e infecções e para explicar os padrões de deterioração e descoloração em árvores feridas, meus colegas e eu desenvolvemos um modelo que chamamos de CODIT (um acrônimo para Compartimentação da Deterioração em Árvores). A primeira parte do modelo representa as respostas que a árvore dá no momento do ferimento. Basicamente, ela reforça as paredes que já estão presentes no lenho, pelo menos, em grande parte. A parede 1 resiste à propagação vertical da madeira infectada; a parede 2, à propagação na direção interior; e a parede 3, à propagação lateral. A segunda parte do modelo representa uma resposta que a árvore dá mais tarde. Basicamente, ela cria uma parede que não estava presente no momento da lesão: o câmbio gera a parede 4, que separa a madeira infectada da madeira recém-formada e saudável.
A parede 1 não existe realmente como uma entidade anatômica antes da infecção. Ela é principalmente uma parede de obstrução, que resiste à propagação vertical da infecção por meios anatômicos e químicos. A árvore possui canalização vertical, constituída, como descrevi acima, de vasos em madeira duras e traqueias em coníferas. Na sequência de uma infecção, os tubos devem ser tampados; afinal, eles são uma maneira fácil para os invasores se espalharem. As árvores fazem isso de várias maneiras. As células ao redor os tubos (chamados de parênquima de contato) podem inflar, obstruindo-os. Pode-se formar incrustações nas suas extremidades. O material granular ou cristalino pode enchê-los. As cavidades entre os tubos podem se fechar. Às vezes, se desenvolvem bolhas de ar, impedindo o transporte de líquidos. A defesa da parede 1 é relativamente fraca, pois, de outra forma, a árvore não poderia sobreviver se obstruísse toda a sua canalização vertical. Além disso, a propagação vertical da infecção é relativamente insignificante: o cerne inteiro de uma árvore pode ser infectado, mas as novas árvores formadas pelo câmbio nos anos subsequentes manterão ela viva.
A parede 2 e a parede 3 já existiam antes do ferimento: elas são formados pelos anéis anuais, que resistem à propagação interna da infecção, e pelos raios parenquimatosos, que resistem à propagação lateral. Após a infecção, as paredes são reforçadas quimicamente. Em parte, no entanto, a parede 2 é anatômica. Em algumas árvores, como o plátano (mas não, por exemplo, o olmo), o final de cada estação de crescimento é marcado pela produção, por parte do câmbio, de uma camada final do que se chama parênquima marginal ou terminal em madeiras duras e células de lenho tardio de paredes grossas em coníferas. As células formam uma barreira celular robusta no perímetro de cada anel de crescimento. A parede 2 é moderadamente forte; a parede 3 é a mais forte das três. Se a última falhar, a deterioração pode se espalhar rapidamente. Esta é a causa das árvores ocas, usadas para proteção e moradia pelo ser humano e pelos animais. A árvore em si pode sobreviver, com copas cheias de folhas saudáveis, devido ao trabalho do câmbio em épocas de crescimento subsequentes à lesão.
A resistência crucial à infecção é aquela que impede sua propagação de dentro para fora – especialmente a defesa do câmbio contra a deterioração que vem do interior da árvore. Aqui entra em jogo a parede 4, a parede do câmbio. Ela é bastante fraca no sentido de fortalecer a estrutura da árvore, mas bastante forte no sentido de ser uma barreira contra microrganismos, que isola o tecido que está fora da lesão e que, portanto, se forma após ela ter acontecido. De fato, é impermeável à maioria dos fungos e bactérias que habitam a madeira ou a casca da árvore. O trabalho recente de R. B. Pearce, P. J . Holloway e Jill Rutherford da Universidade de Oxford estabelece que as células de barreira da parede 4 são revestidas com suberina, um ácido graxo que dá à casca externa sua resistência aos invasores microbianos (eles quase nunca possuem enzimas capazes de atuar sobre a suberina).
Meus colegas e eu também inventamos um dispositivo que detecta madeira deteriorada em árvores vivas sem danificá-las. O dispositivo se aproveita de uma circunstância da deterioração: à medida que a madeira se decompõe, seu conteúdo de portadores de carga elétrica (principalmente íons de potássio) aumenta, de modo que a resistência elétrica do tecido, medida em ohms, diminui. Nosso equipamento, portanto, consiste em um gerador à bateria de corrente pulsada, uma sonda e um ohmímetro. Para testar a deterioração, um furo de 0,24 cm (3/32”) de diâmetro é realizado com 20 a 30cm (8 a 12”) de profundidade na árvore. A sonda é inserida lentamente. Uma diminuição da resistência elétrica encontrada na ponta da sonda (a ponta não tem isolamento) sinaliza o apodrecimento. O dispositivo também está sendo empregado para detectar a deterioração na linha do chão em postes enterrados.
Meus colegas e eu estamos agora colaborando com geneticistas num esforço para descobrir quais indivíduos dentro de uma determinada espécie de árvores têm a melhor capacidade de compartimentação. (Essa capacidade é controlada fortemente pela genética.) Amparados com nossos novos conceitos de biologia das árvores, estamos também reexaminando suas doenças. Descobrimos, por exemplo, que os olmos compartimentam a madeira infectada pelo fungo que causa a doença do olmeiro holandês. A defesa pode levar à inanição à medida que os tecidos que normalmente armazenam energia são murados. Além disso, estamos planejando correções em muitos dos procedimentos padrão de cuidado de árvores, tais como a poda.
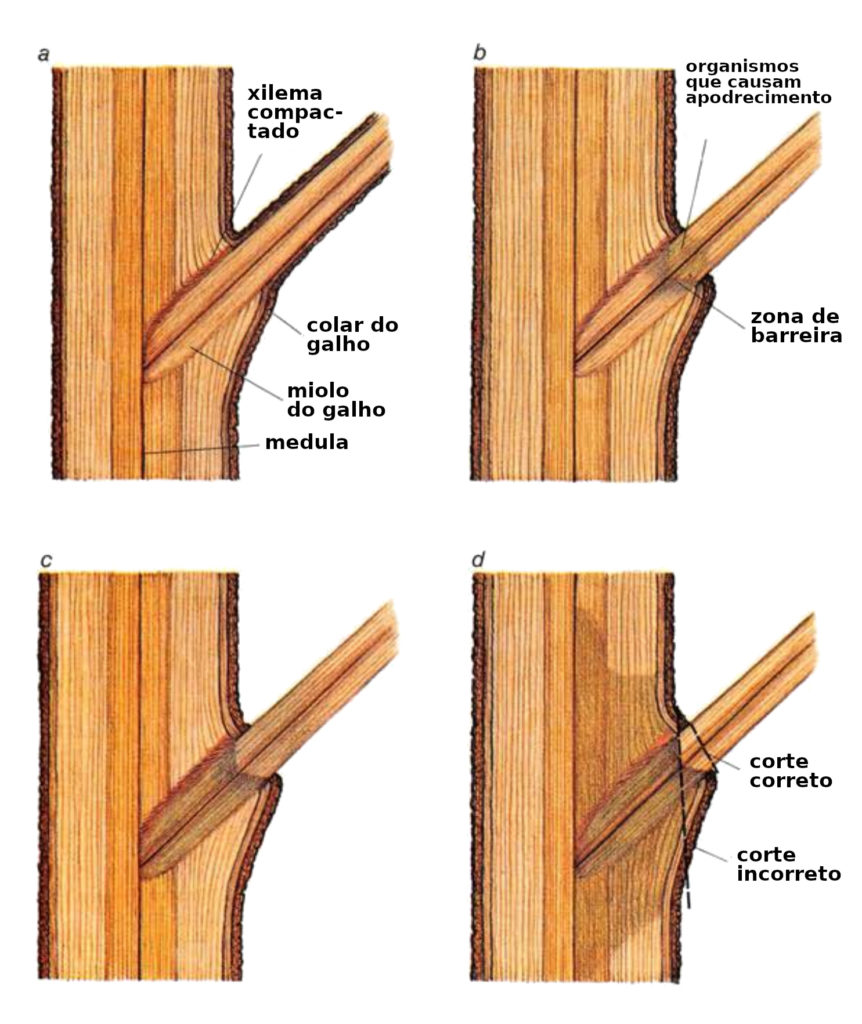
Muito do mal-entendido que está por trás dos cuidados impróprios com as árvores acontece porque conceitos desenvolvidos para explicar a biologia animal são aplicados, quase inconscientemente, às árvores. Várias vezes, elas são tratadas como animais ou, pior ainda, como pessoas. Os curativos são colocados em árvores feridas num esforço para parar a deterioração e promover a cura, assim como um adulto cobre um corte em uma criança. As cavidades de podridão são limpas além da parte que está podre, expondo a madeira saudável, da mesma forma que um dentista limpa um dente podre. Os galhos são podados diretamente no tronco, e em alguns países, é feita uma incisão em forma de diamante na casca logo abaixo do galho; o calo subsequente, ou cicatriz, é considerado um sinal de que a árvore está se curando bem.
Nenhum desses tratamentos é benéfico para as árvores; de fato, todos eles podem ser prejudiciais. Nenhum dado científico mostra que qualquer substância aplicada a uma ferida em uma árvore irá ajudá-la a parar de se decompor. Os curativos são principalmente cosméticos. (Talvez, isso funcione mais como um remédio psicológico para a pessoa dona da árvore.) A limpeza de uma cavidade para além da parte podre, adentrando na madeira saudável, promove um apodrecimento ainda maior. De fato, é uma das piores coisas que se pode fazer a uma árvore. A cavidade existe porque a árvore se murou contra o apodrecimento. Finalmente, a poda inadequada permite que o apodrecimento tome conta das células feridas no tronco. Ao redor da base de cada galho há um inchaço conhecido como colar. Dentro dele, está a zona de proteção do galho, ou seja, o lugar onde as defesas químicas do ramo são estabelecidas. O colar não deve ser ferido, muito menos removido, pela poda.
A deterioração nas árvores é um processo natural. Em alguns casos, pode ser controlado: acelerando, retardando ou estagnando-o. Quando não pode ser influenciado, pode pelo menos ser detectado, de forma não destrutiva, em árvores vivas. Além disso, pode ser previsto, tanto na sua taxa de propagação como na sua configuração. Além disso, as árvores podem ser selecionadas por sua resistência à propagação de deterioração. A silvicultura científica está se aproximando de uma nova compreensão das árvores e de como elas sobrevivem sob muitas pressões.